
описание с инструкцией по замене
Независимый автомобильный портал «Авто Центр Ру» / Датчик холостого и заднего хода Lada Kalina: описание с инструкцией по замене
Для стабильной работы двигателя вхолостую предназначен датчик холостого хода Калина. В данной статье речь пойдет о Ладе Калина – о характерных признаках прибора на этом автомобиле, о том, какие могут возникнуть неполадки и как их устранить, а также будут даны рекомендации по замене устройства.
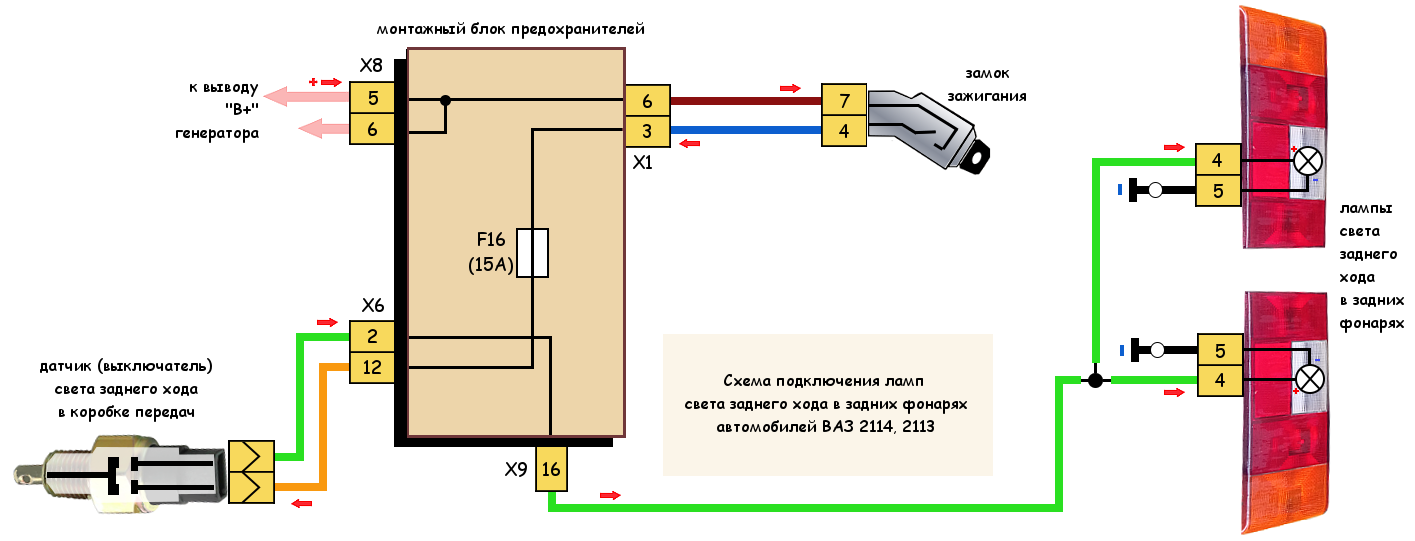
Датчик заднего хода
Характеристика контроллера ХХ
Давайте ознакомимся подробнее с датчиком ХХ.
Назначение и принцип работы
Датчик холостого хода – это электродвигатель с конусной иглой определенной толщины. Это устройство расположено на корпусе дроссельного узла рядом с устройством положения дроссельной заслонки, и крепится двумя винтиками или при помощи технического лака.
Приспособление имеет единственную цель: стабилизировать обороты двигателя вхолостую, и происходит все таким образом:
- В двигатель, работающий вхолостую, поступает некоторый объем воздуха, который позволяет ему нормально работать.

- Прибор не считывает и не контролирует ничего, он выполняет то что ему дает ЭБУ. Блок контролирует, считывает и согласно показаниям тахометра дает команду на регулятор ХХ .
- Если прогреть двигатель автомобиля до оптимальной температуры, контроллер будет поддерживать обороты. Если он не прогретый, то за счет работы РХХ контроллер увеличит обороты. Эта манипуляция обеспечит на повышенных оборотах оптимальную температуру двигателя, анализируя работу (автор — RussianTuning).

Типичные неисправности
Как и у любого систематически работающего устройства, у ДХХ могут возникать различные неполадки, например:
- Машина на холостом внезапно глохнет.
- Обороты ДХХ не систематические и «плавают».
- При запуске двигателя без предварительного прогрева контроллер не начинает работу и обороты датчика не совершаются так, как должны.

- Внезапно глохнет при переключении передачи на коробке передач.
- Обороты ДХХ виснут после отпускания педали гада или при активации различной автомобильной техники.
Когда с автомобилем происходят все эти странные вещи, очевидно, что проблема заключается в регуляторе. К сожалению, самодиагностики этого прибора нет, поэтому чтобы разобраться, в чем именно причина неисправности, нужно проверить его самостоятельно. И если починить не удастся, то заменить. Замена датчика холостого хода на Калине будет освещена ниже.
Месторасположение детали
Руководство по замене датчика ХХ
Для замены датчика холостого хода не потребуется много времени и наличия особых навыков.
- Необходимо заглушить двигатель и дать автомобилю немного остыть.
- Отключить аккумулятор.
- Изъять из дроссельного узла нужный регулятор.
- Отсоединить штекер и выкрутить два болта. Затем вытащить полностью конструкцию.
- Вернуть с новым датчиком все в прежнее положение в обратной схеме.

Фотогалерея «Инструкция по монтажу»
Ниже можно узнать, каким образом происходит демонтаж.
1. Найти место регулятора.
2. Изъять регулятор из дроссельного узла.
3. Вытащить датчик холостого хода.
Характеристика датчика ЗХ
Давайте разбираться, что же представляет собой контроллер ЗХ!
Назначение и принцип работы
Датчик заднего хода на Лада Калина находится на коробке передач. Цель этого устройства заключается в том, чтобы задние фары выключались сразу же при прекращении заднего хода. На упомянутой выше коробке передач установлена специальная катушка, которая занимается тем, то блокирует заднюю передачу при возникновении проблем с датчиком.
На рычаге коробки есть кольцо, при помощи которого дается задний ход. Едва эта специальная деталь приподнимается, на катушку дается заряд тока, который втягивает сердечник катушки. Благодаря этим действиям освобождается место для переключения задней передачи. Этот импульс подает также сигнал о включении индикаторов заднего хода. В случае выключения фар создается обратная ситуация, когда сердечник становится на место.
Этот импульс подает также сигнал о включении индикаторов заднего хода. В случае выключения фар создается обратная ситуация, когда сердечник становится на место.
Месторасположение детали заднего хода
Типичные неисправности
Лада Калина неплохая машина, но есть в ней одна особенность, которая на корню портит многие достоинства. Эта особенность касается включения заднего хода. Проводки на коробке передач столь тонкие и уязвимые, что порваться могут в любую минуту, если не обращаться с ними аккуратно.
Есть и другие часто случающиеся неполадки:
- Когда можно дать задний ход, но лампа не горит. Если речь идет об одной лампе, значит дело в ней и потребуется ее замена, если не горят обе – нужно исследовать цепь между деталью и лампами.
- Проблема цепи индексации заднего хода. В таком случае нужно диагностировать кольцо на коробке передач.
- Часто работа прекращается из-за проводков, которые ведут от датчика на дно машины, где на провод влияет окружающая среда и контакт запросто может окислиться.

Чистка дроссельного узла
Руководство по замене регулятора заднего хода
Чтобы произвести замену, даже обычному автолюбителю потребуется наличие некоторых навыков и знаний.
- Прежде всего ЗХ, находящийся под машиной, нужно отсоединить от проводки.
- Выключатель нужно осторожно изъять из коробки и мгновенно заменить его новым, чтобы не вытекло масло. Под коробкой стоять ни в коем случае нельзя, ведь масло может попасть на лицо и одежду.
- Лучше всего, чтобы перед заменой машина немного остыла, ведь в таком случае давление на масло не будет обильным.
Видео «Замена датчика фаз Лада Калина»
Подробнее о том, как заменить устройство на своем автомобиле, можно узнать из видео от автора Alexandr V.
Извините, в настоящее время нет доступных опросов.
На Калине не включается задняя передача: возможные причины
Автовладельцы нередко сталкиваются с ситуацией, когда на «Калине» не включается задняя передача. Причины этой неполадки нередко не могут определить даже на СТО. Но если взглянуть в сервисный мануал и почитать информацию по этому автомобилю, можно узнать, в чем причина и как устранить данную неполадку.
Причины этой неполадки нередко не могут определить даже на СТО. Но если взглянуть в сервисный мануал и почитать информацию по этому автомобилю, можно узнать, в чем причина и как устранить данную неполадку.
На самом деле, устраняется эта проблема достаточно просто. Нет необходимости в серьезном ремонте, требующем демонтажа КПП (за исключением отдельных случаев). Чаще всего всему виной электронная блокировка, проводка и предохранитель.
Трансмиссия
Это самая первая причина. К не включению задней скорости часто приводит износ узлов – передача вовсе не будет включаться или ее выбивает на ходу. Первое, на что следует обратить внимание, – это масло. При потере физических и химических свойств вязкость его значительно меняется. Это может привести к заклиниванию передач. Механизм коробки будет работать, но с включением определенных скоростей могут возникнуть определенные сложности. Вторая причина, связанная с механизмом КПП, – естественный износ внутренних элементов трансмиссии. Так, если не включается задняя передач на «Калине», то «подозреваемых» несколько:
Так, если не включается задняя передач на «Калине», то «подозреваемых» несколько:
- Шестеренка задней передачи.
- Выжимной подшипник.
- Вилка включения.
Коробку вскрывают, проводят дефектовку, изношенную деталь заменяют. После этого механизм КПП должен работать исправно.
Система сцепления
Еще одной популярной причиной подобной проблемы является сцепление. Диагностировать данную неисправность довольно легко. В момент отпускания педали передача не будет заходить, либо включаться с характерным хрустом. Это не говорит о поломке задней передаче «Лады Калины» — это дефект сцепления.
Так, стоит обратить внимание на выжимной подшипник, а также на диск. При значительной выработке эти детали следует заменить.
Электрика
Это последняя поломка, из-за которой не включается задний ход на «Ладе Калине». В автомобиле имеется система электронной блокировки заднего хода. Данная система очень полезная, но нередко выходит из строя. Почему так случается? Иногда перегорает предохранитель. В колодке он под номером F21. Также следует проверить штекер трансмиссии под капотом автомобиля.
В колодке он под номером F21. Также следует проверить штекер трансмиссии под капотом автомобиля.
Короткоходная кулиса на автомобили ВАЗ-Приора
Для более четкого переключения коробки передач служит короткоходная кулиса с секвентальной…
Предохранитель найти достаточно легко – он находится в блоке (восьмой слева). Его рекомендуется для проверки извлечь. Проверять лучше мультиметром.
Салонные провода
Когда предохранитель полностью исправен, со штекером все в порядке, а на «Калине» не включается задняя передача, смотрят выключатель, находящийся под декоративным чехлом около самого рычага КПП. К этому выключателю подведены два провода. Их замыкают между собой. Если передача начала нормально включаться, то причина была именно в выключателе. Можно измерить напряжение на проводах при помощи иголок. На выключателе должно быть не менее 12 В. При отсутствии мультиметра измерить напряжение можно при помощи лампочки. Если она не горит, это свидетельствует о переломе провода. Обычно подобное случается в том месте, где устанавливается хомут на поддоне. Это легко объясняет вопрос: почему не включается задняя передача на Ладе Калине.
Обычно подобное случается в том месте, где устанавливается хомут на поддоне. Это легко объясняет вопрос: почему не включается задняя передача на Ладе Калине.
Киста матки: возможные причины, симптомы, методы…
Сегодня доброкачественные новообразования часто встречаются в гинекологии, они диагностируются у 15…
Хомут снимают и сжимают в нижней части крепежа. Далее отсоединяют разъем выключателя ламп. Затем достают весь жгут. Переломанный провод будет виден сразу же. Что делать далее? Шнур соединяют с отрезком хорошего провода – можно сделать простую скрутку. Но лучше всего пропаять соединение. Далее все собирается в обратном порядке. Если передача включается, то проблема решена.
Соленоид
Это последняя причина, из-за которой на «Калине» не включается задняя передача. Она связана с соленоидом. Для его проверки лучше использовать мультиметр. Но если его нет, то в отверстие для подвода тока вставляют булавки. Наличие напряжения проверяют обычной 12-вольтовой лампочкой.
Специалисты утверждают, что проблему можно победить с помощью ключа «на 22». Система блокировки включения задней скорости на «Калине» реализована при помощи этого соленоида. Питание на него подается при помощи выключателя в виде кольца на селекторе КПП. Когда кольцо поднимается, шток соленоида втягивается внутрь и блокировка отключается. Вместе с этим срабатывает соответствующий концевой выключатель, который включает лампы заднего хода и «пищалку» в салоне. Через предохранитель на соленоид от аккумулятора приходит 12 В. С помощью выключателя, расположенного на селекторе, осуществляется замыкание на массу. Далее срабатывает катушка на соленоиде.
Как отремонтировать соленоид?
Когда на «Калине» не включается задняя передача, можно попробовать восстановить соленоид. Где он находится? Если вывернуть рулевое колесо вправо, а затем заглянуть под автомобиль от левого колеса, можно увидеть соленоид. Он находится на КПП. К соленоиду подсоединен разъем и провода. Разъем снимают, а ключом «на 22» откручивают соленоид приблизительно на восемь миллиметров. В получившийся зазор подматывают провод или же изолирующую ленту. Затем снова затягивают соленоид. Далее можно пробовать включить заднюю передачу. При движении на автомобиле стоит быть максимально внимательным – первая передача находится рядом.
В получившийся зазор подматывают провод или же изолирующую ленту. Затем снова затягивают соленоид. Далее можно пробовать включить заднюю передачу. При движении на автомобиле стоит быть максимально внимательным – первая передача находится рядом.
Обратите внимание! В случае замены соленоида придется заменить масло в трансмиссии. Но если успеть заткнуть отверстие, то достаточно будет просто долить нужное количество смазывающей жидкости. Специалисты рекомендуют использовать масло «Тотал Трансмишн» вязкостью 75W80. Необходимый объем – 2,2 литра для коробки 2180 и 3,3 литра для 2181 (с тросиковым приводом).
Если проверки ни к чему не привели
Бывает и так, что диагностика ничего не дала – ни одна из описанных причин не подходит. В этом случае есть три возможных варианта:
- Вышел из строя механизм выбора передач.
- Срезало фиксирующий болт.
- Лопнула пружина.
Самая страшное в этой ситуации, что любая из этих причин требует снятия коробки. Это уже вполне серьезный ремонт, который под силу далеко не всем.
Это уже вполне серьезный ремонт, который под силу далеко не всем.
Заключение
Это все причины, по которым не включается задняя передача на «Калине». Первым делом следует продиагностировать электрику, выключатель, а также соленоид. Затем уже переходят к демонтажу и разборке механизма КПП и сцепления с заменой всех износившихся узлов.
Viburnum opulus Фенольные соединения плодов как цитозащитные агенты, способные снижать поглощение свободных жирных кислот и глюкозы клетками Caco-2
1. Cory H., Passarelli S., Szeto J., Tamez M., Mattei J. The Role of Polyphenols в области здравоохранения и продовольственных систем: мини-обзор. Передний. Нутр. 2018;5:87. doi: 10.3389/fnut.2018.00087. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2. Экен А., Юджел О., Ипек И., Айше Б., Эндирлик Б.У. Исследование защитного эффекта Viburnum opulus L. Экстракт плодов против окислительного стресса, вызванного ишемией/реперфузией, после трансплантации легких у крыс. Kafkas Üniversitesi Veteriner Fakültesi Dergisi. 2017; 23:437–444. [Google Scholar]
Kafkas Üniversitesi Veteriner Fakültesi Dergisi. 2017; 23:437–444. [Google Scholar]
3. Перова И.Б., Жогова А.А., Черкашин А.В., Эллер К.И., Раменская Г.В. Биологически активные вещества из плодов калины европейской. фарм. хим. Дж. 2014; 48:332–339. doi: 10.1007/s11094-014-1105-8. [CrossRef] [Google Scholar]
4. Молдован Б., Дэвид Л., Чишбора К., Чимпою К. Кинетика деградации антоцианов из европейской клюквы ( Viburnum opulus L.) Фруктовые экстракты. Влияние температуры, pH и хранения растворителя. Молекулы. 2012;17:11655–11666. doi: 10.3390/молекулы171011655. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
5. Кызылай О.Н., Юлкер Ф., Челик В., Оздемир Т., Чакмак О., Джан Э. Оценка эффективности Гилабуру ( Экстракт Viburnum opulus L. в медикаментозном экспульсивном лечении камней дистальных отделов мочеточника Оценка эффективности Gilaburu ( Viburnum opulus L.) при медикаментозном экспульсивном лечении камней дистального отдела мочеточника. Турок. Дж. Урол. 2019 г.: 10.5152/tud.2019.23463. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Турок. Дж. Урол. 2019 г.: 10.5152/tud.2019.23463. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Джейлан Д., Аксой А., Эртекин Т., Яй А.Х., Нисари М., Каратопрак Г.Ш., Ульгер Х. Эффекты гилабуру (Viburnum opulus) при экспериментально индуцированной асцитной опухоли Эрлиха у мышей. Дж. Рак Рез. тер. 2018;14:310–320. [PubMed] [Google Scholar]
7. Козловская В., Вагнер К., Мур Е.М., Матковский А. Ботаническое происхождение традиционных лекарственных средств из Карпат на украинско-польской границе. Передний. Фармакол. 2018;9:295. doi: 10.3389/fphar.2018.00295. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Заклош-Шида М., Маевска И., Редзыня М., Козиолкевич М. Антидиабетический эффект полифенольных экстрактов из выбранных пищевых растений в виде α-амилазы, Ингибиторы α-глюкозидазы и PTP1B и цитопротекторы β-клеток поджелудочной железы — сравнительное исследование. Курс. Вершина. Мед. хим. 2015;15:2431–2444. doi: 10.2174/1568026615666150619143051. [PubMed] [CrossRef] [Google Scholar]
[PubMed] [CrossRef] [Google Scholar]
9. Михайлова Д., Попова А., Алексиева И., Красстанов А., Ланте А. Полифенолы как подходящие средства контроля ожирения и диабета. Откройте Биотех. Дж. 2018; 12:219–228. doi: 10.2174/1874070701812010219. [CrossRef] [Google Scholar]
10. Фаррелл Т.Л., Эллам С.Л., Форрелли Т., Уильямсон Г. Ослабление транспорта глюкозы через монослои клеток Caco-2 с помощью богатого полифенолами растительного экстракта: взаимодействие с транспортерами SGLT1 и GLUT2. Биофакторы. 2013; 39: 448–456. doi: 10.1002/biof.1090. [PubMed] [CrossRef] [Google Scholar]
11. Kwon O., Eck P., Chen S., Corpe CP, Lee JH, Kruhlak M., Levine M. Ингибирование кишечного транспортера глюкозы GLUT2 флавоноидами. FASEB J. 2007; 21: 366–377. дои: 10.1096/fj.06-6620com. [PubMed] [CrossRef] [Google Scholar]
12. Шрек К., Мельциг М.Ф. Кишечные транспортеры насыщенных длинноцепочечных жирных кислот, глюкозы и фруктозы и их ингибирование натуральными растительными экстрактами в клетках Caco-2. Молекулы. 2018;23:2544. doi: 10.3390/молекулы23102544. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Молекулы. 2018;23:2544. doi: 10.3390/молекулы23102544. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Manzano S., Williamson G. Полифенолы и фенольные кислоты из клубники и яблока снижают поглощение и транспорт глюкозы клетками Caco-2 кишечника человека. Мол. Нутр. Еда Рез. 2010;54:1773–1780. doi: 10.1002/mnfr.201000019. [PubMed] [CrossRef] [Google Scholar]
14. Stremmel W., Staffer S., Wannhoff A., Pathil A. Общее всасывание жирных кислот, контролируемое базолатеральной экскрецией хиломикронов при регуляции p-JNK1. Биохим. Биофиз. Акта-мол. Клеточная биол. Липиды. 2017; 1862: 917–928. doi: 10.1016/j.bbalip.2017.05.013. [PubMed] [CrossRef] [Google Scholar]
15. Podsędek M., Majewska A., Redzynia I., Sosnowska M., Koziołkiewicz D. Ингибирующее действие in vitro на пищеварительные ферменты и антиоксидантный потенциал обычно потребляемых фруктов. Дж. Агрик. Пищевая хим. 2014;62:4610–4617. doi: 10.1021/jf5008264. [PubMed] [CrossRef] [Академия Google]
16. Будрин Г., Заклош-Шида М., Зачиньска Д., Жижелевич Д., Гжельчик Ю., Здуньчик З., Юшкевич Ю. Экстракты зеленого и обжаренного кофе как антиоксиданты в клетках βTC3 при индуцированном окислительном стрессе и накоплении липидов ингибиторы в клетках 3T3L1 и их биоактивность у крыс, получавших диету с высоким содержанием жиров. Евро. Еда Рез. Технол. 2017; 243:1323–1334. doi: 10.1007/s00217-017-2843-0. [CrossRef] [Google Scholar]
Будрин Г., Заклош-Шида М., Зачиньска Д., Жижелевич Д., Гжельчик Ю., Здуньчик З., Юшкевич Ю. Экстракты зеленого и обжаренного кофе как антиоксиданты в клетках βTC3 при индуцированном окислительном стрессе и накоплении липидов ингибиторы в клетках 3T3L1 и их биоактивность у крыс, получавших диету с высоким содержанием жиров. Евро. Еда Рез. Технол. 2017; 243:1323–1334. doi: 10.1007/s00217-017-2843-0. [CrossRef] [Google Scholar]
17. Заклош-Шида М., Павлик Н. Айва японская ( Chaenomeles japonica L.) полифенольный экстракт фруктов модулирует углеводный обмен в клетках HepG2 посредством AMP-активируемой протеинкиназы. Акта Биохим. пол. 2018;65:67–78. doi: 10.18388/abp.2017_1604. [PubMed] [CrossRef] [Google Scholar]
18. Хоган А.М., Сваминатан В., Паллегар Н.К., Кристиан С.Л. Нильский красный и 2-НБДГ несовместимы для одновременного определения накопления липидов и глюкозы. Междунар. Дж. Спектроск. 2016;2016:5215086. doi: 10.1155/2016/5215086. [CrossRef] [Академия Google]
19. Новак А., Клевицкий Р., Липин Л. Эллагитаннины из Rubus idaeus L. Оказывают гено- и цитотоксическое действие на клеточную линию аденокарциномы толстой кишки человека Caco-2. Дж. Агрик. Пищевая хим. 2017;65:2947–2955. doi: 10.1021/acs.jafc.6b05387. [PubMed] [CrossRef] [Google Scholar]
Новак А., Клевицкий Р., Липин Л. Эллагитаннины из Rubus idaeus L. Оказывают гено- и цитотоксическое действие на клеточную линию аденокарциномы толстой кишки человека Caco-2. Дж. Агрик. Пищевая хим. 2017;65:2947–2955. doi: 10.1021/acs.jafc.6b05387. [PubMed] [CrossRef] [Google Scholar]
20. Peng BJ, Zhu Q., Zhong Y.L., Xu S.H., Wang Z. Хлорогеновая кислота поддерживает гомеостаз глюкозы посредством модуляции экспрессии SGLT-1, GLUT-2 и PLG. в различных сегментах кишечника крыс Sprague-Dawley, получавших диету с высоким содержанием жиров. Биомед. Окружающая среда. науч. 2015;28:894–903. [PubMed] [Google Scholar]
21. Велиоглу Ю.С., Экиджи Л., Пойразоглу Э.С. Оригинальная статья Фенольный состав ягод клюквы европейской ( Viburnum opulus L.) и устранение терпкости ее товарного сока. Междунар. Дж. Пищевая наука. Технол. 2006;9205:1011–1015. doi: 10.1111/j.1365-2621.2006.01142.x. [CrossRef] [Google Scholar]
22. Карачелик П., Кучук А.А., Искефиели М. , Айдемир З., де Смет С., Мисерез С., Сандра Б. Антиоксидантные компоненты Viburnum opulus л. определено методами он-лайн ВЭЖХ-УФ-ABTS для удаления радикалов и методами ЖХ-УФ-ESI-MS. Пищевая хим. 2015; 175:106–114. doi: 10.1016/j.foodchem.2014.11.085. [PubMed] [CrossRef] [Google Scholar]
, Айдемир З., де Смет С., Мисерез С., Сандра Б. Антиоксидантные компоненты Viburnum opulus л. определено методами он-лайн ВЭЖХ-УФ-ABTS для удаления радикалов и методами ЖХ-УФ-ESI-MS. Пищевая хим. 2015; 175:106–114. doi: 10.1016/j.foodchem.2014.11.085. [PubMed] [CrossRef] [Google Scholar]
23. Kraujalyte V., Rimantas P., Pukalskas A., Laima C. Антиоксидантные свойства и полифенольный состав плодов разных генотипов клюквы европейской ( Viburnum opulus L.). Пищевая хим. 2013; 141:3695–3702. doi: 10.1016/j.foodchem.2013.06.054. [PubMed] [CrossRef] [Академия Google]
24. Де Фрейтас В., Фариа А., Пестана Д., Азеведо Дж., Азеведо И., Матеус Н. Поглощение антоцианов эпителиальными клетками кишечника — предполагаемое участие GLUT2. Мол. Нутр. Еда Рез. 2009;53:1430–1437. [PubMed] [Google Scholar]
25. Hajiaghaalipour F., Khalilpourfarshbafi M., Arya A. Модуляция белка-переносчика глюкозы диетическими флавоноидами при сахарном диабете 2 типа. Междунар. Дж. Биол. науч. 2015;11:508. doi: 10.7150/ijbs.11241. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Биол. науч. 2015;11:508. doi: 10.7150/ijbs.11241. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Catarino T.A., Gonc P. Влияние окислительного стресса на поглощение бутирата кишечным эпителием. Евро. Дж. Фармакол. 2013; 699: 88–100. [PubMed] [Google Scholar]
27. Маргина Д., Градинару Д., Манда Г., Неаго И., Илие М. Мембранарные эффекты, оказываемые in vitro полифенолами — кверцетин, эпигаллокатехин галлат и куркумин — на клетки HUVEC и Jurkat. , актуально при сахарном диабете. Пищевая хим. Токсикол. 2013;61:86–93. doi: 10.1016/j.fct.2013.02.046. [PubMed] [CrossRef] [Академия Google]
28. Амараратна М., Джонстон М.Р., Рупасингх Х.П.В. Растительные полифенолы как химиопрофилактические средства против рака легких. Междунар. Дж. Мол. науч. 2016;17:1352. doi: 10.3390/ijms17081352. [Статья PMC бесплатно] [PubMed] [CrossRef] [Google Scholar]
29. Goszcz K., Duthie G.G., Stewart D., Leslie S.J., Megson I.L. Биоактивные полифенолы и сердечно-сосудистые заболевания: химические антагонисты, фармакологические агенты или ксенобиотики, которые вызывают адаптивный ответ? бр. Дж. Фармакол. 2017; 174:1209–1225. doi: 10.1111/bph.13708. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Фармакол. 2017; 174:1209–1225. doi: 10.1111/bph.13708. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Лян Н., Дюпюи Дж.Х., Яда Р.Ю., Киттс Д.Д. Изомеры хлорогеновой кислоты напрямую взаимодействуют с передачей сигналов Keap 1-Nrf2 в клетках Caco-2. Мол. Клетка. Биохим. 2019; 457:105–118. doi: 10.1007/s11010-019-03516-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Bonarska-kujawa H.P.D. Влияние хлорогеновой кислоты на фазовый переход в фосфолипидных и фосфолипидно-холестериновых мембранах. J. Термальный анализ. Калорим. 2014; 118:943–950. [Google Scholar]
32. Лян Н., Киттс Д.Д. Изомеры хлорогеновой кислоты (CGA) уменьшают выработку интерлейкина 8 (IL-8) в клетках Caco-2 за счет снижения фосфорилирования p38 и повышения целостности клеток. Междунар. Дж. Мол. науч. 2018;19:3837. doi: 10.3390/ijms19123873. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Ван Дж., Ли Дж., Лю Дж., Сюй М., Тонг С., Ван Дж. Хлорогеновая кислота предотвращает повреждение ДНК, вызванное изопротеренолом. в гладкомышечных клетках сосудов. Мол. Мед. Отчет 2016; 14: 4063–4068. doi: 10.3892/mmr.2016.5743. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Хлорогеновая кислота предотвращает повреждение ДНК, вызванное изопротеренолом. в гладкомышечных клетках сосудов. Мол. Мед. Отчет 2016; 14: 4063–4068. doi: 10.3892/mmr.2016.5743. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
34. Экбатан С.С., Ли С., Горбани М., Азади Б. Хлорогеновая кислота и ее микробные метаболиты оказывают антипролиферативное действие, клетка S-фазы Задержка цикла и апоптоз в клетках Caco-2 рака толстой кишки человека. Междунар. Дж. Мол. науч. 2018;19:723. doi: 10.3390/ijms123. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Cheah K.Y., Howarth G.S., Bindon K.A., Kennedy J.A., Bastian S.E.P. Низкомолекулярные процианидины из виноградных косточек усиливают воздействие химиотерапии 5-фторурацилом на клетки рака толстой кишки человека Caco-2. ПЛОС ОДИН. 2014;9:e98921. doi: 10.1371/journal.pone.0098921. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
36. Gorlach S., Wagner W., Podsędek A., Szewczyk K., Koziołkiewicz M. , Dastych J. Процианидины из айвы японской (Chaenomeles japonica) фрукты индуцируют апоптоз в клетках Caco-2 рака толстой кишки человека в степени, зависящей от полимеризации. Нутр. Рак. 2011;63:1348–1360. doi: 10.1080/01635581.2011.608480. [PubMed] [CrossRef] [Академия Google]
, Dastych J. Процианидины из айвы японской (Chaenomeles japonica) фрукты индуцируют апоптоз в клетках Caco-2 рака толстой кишки человека в степени, зависящей от полимеризации. Нутр. Рак. 2011;63:1348–1360. doi: 10.1080/01635581.2011.608480. [PubMed] [CrossRef] [Академия Google]
37. Родригес-Рамиро И., Рамос С., Браво Л., Гойя Л., Мартин М.А. Процианидин B2 и полифенольный экстракт какао ингибируют вызванный акриламидом апоптоз в клетках Caco-2 человека, предотвращая окислительный стресс и активируя путь JNK. Дж. Нутр. Биохим. 2011; 22:1186–1194. doi: 10.1016/j.jnutbio.2010.10.005. [PubMed] [CrossRef] [Google Scholar]
38. Пандей М.К., Гупта С.К., Набавизаде А., Аггарвал Б.Б. Семинары по биологии рака Регулирование сигнальных путей клеток диетическими агентами для профилактики и лечения рака. Семин. Рак биол. 2017; 46: 158–181. doi: 10.1016/j.semcancer.2017.07.002. [PubMed] [CrossRef] [Академия Google]
39. Дурия Ю.К., Шарма Д. Некроптоз: регулируемый воспалительный способ гибели клеток. Дж. Нейровоспаление. 2018;15:199. doi: 10.1186/s12974-018-1235-0. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Нейровоспаление. 2018;15:199. doi: 10.1186/s12974-018-1235-0. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
40. Lin W., Tongyi S. Роль членов семейства Bax/Bcl-2 в индуцированном полифенолами зеленого чая некроптозе p53-дефицитных клеток Hep3B. Опухоль биол. 2014; 35:8065–8075. doi: 10.1007/s13277-014-2064-0. [PubMed] [CrossRef] [Google Scholar]
41. Фам Т., Климченко А., Колло М. Последние достижения в области флуоресцентных зондов для липидных капель. Материалы. 2018;11:1768. дои: 10.3390/ma11091768. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Rohm B., Riedel A., Ley J.P., Widder S. Активация функции и увеличение ацетил-коэнзима A. Food Funct. 2015;6:173–185. [PubMed] [Google Scholar]
43. Glatz J.F.C., Luiken J.J.F.P. Биохимия От жира к жиру (CD36/SR-B2): понимание регуляции поглощения жирных кислот клетками. Биохимия. 2017; 136:21–26. doi: 10.1016/j.biochi.2016.12.007. [PubMed] [CrossRef] [Google Scholar]
44. Schneider H., Staudacher S., Poppelreuther M., Stremmel W., Ehehalt R., Füllekrug J. Белок-опосредованное поглощение жирных кислот: синергизм между CD36/FAT-облегченными транспорт и метаболизм, управляемый ацил-КоА-синтетазой. Арка Биохим. Биофиз. 2014; 546:8–18. doi: 10.1016/j.abb.2014.01.025. [PubMed] [CrossRef] [Академия Google]
Schneider H., Staudacher S., Poppelreuther M., Stremmel W., Ehehalt R., Füllekrug J. Белок-опосредованное поглощение жирных кислот: синергизм между CD36/FAT-облегченными транспорт и метаболизм, управляемый ацил-КоА-синтетазой. Арка Биохим. Биофиз. 2014; 546:8–18. doi: 10.1016/j.abb.2014.01.025. [PubMed] [CrossRef] [Академия Google]
45. Перона Дж.С. Biochimica et Biophysica Acta Мембранные липидные изменения при метаболическом синдроме и роль диетических масел. BBA-Биомембр. 2017; 1859:1690–1703. doi: 10.1016/j.bbamem.2017.04.015. [PubMed] [CrossRef] [Google Scholar]
46. Sancheza M.B., Miranda-Pereza E., Verjanb JCG, Barreraa M.D.F., Perez-Ramosc J., Alarcon-Aguilaret F.J. Потенциал хлорогеновой кислоты как многоцелевого агента: инсулин- стимулятор секреции и двойной агонист PPAR α/γ. Биомед. Фармацевт. 2017;94: 169–175. doi: 10.1016/j.biopha.2017.07.086. [PubMed] [CrossRef] [Google Scholar]
47. Pang Y., Zhu Q., Kang J., Liu M., Wang Z. Хлорогеновая кислота действует как новый агонист PPAR γ 2 во время дифференцировки мышиного 3T3. -L1 Преадипоциты. Биомед Рез. Междунар. 2018 г.: 10.1155/2018/8594767. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
-L1 Преадипоциты. Биомед Рез. Междунар. 2018 г.: 10.1155/2018/8594767. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Vrbacky M. Сукцинимидил олеат, установленный ингибитор транслоказы CD36/FAT, ингибирует комплекс III митохондриальной дыхательной цепи. Биохим. Биофиз. Рез. коммун. 2010;391: 1348–1351. [PubMed] [Google Scholar]
49. Абумрад Н.А., Чифарелли В. Кишечный CD36 и другие ключевые белки утилизации липидов: роль в абсорбции и гомеостазе кишечника. Компр. Физиол. 2018; 8: 493–507. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Zhang D., Zhang R., Liu Y., Sun X., Yin Z., Li H. Варианты гена CD36 связаны с сахарным диабетом 2 типа через взаимодействие ожирения у взрослых сельских жителей Китая. Ген. 2018; 659: 155–159. doi: 10.1016/j.gene.2018.03.060. [PubMed] [CrossRef] [Академия Google]
51. Hemmersbach S., Brauer S.S., Hu S., Galla H., Humpf H. Исследования трансэпителиальной проницаемости флаван-3-ол-C-глюкозидов и димеров и тримеров процианидина через монослой клеток Caco-2. Дж. Агрик. Пищевая хим. 2013;61:7932–7940. doi: 10.1021/jf402019f. [PubMed] [CrossRef] [Google Scholar]
Дж. Агрик. Пищевая хим. 2013;61:7932–7940. doi: 10.1021/jf402019f. [PubMed] [CrossRef] [Google Scholar]
52. Zou T., Feng D., Song G., Li H., Tang H., Ling W. Роль натрий-зависимого транспортера глюкозы 1 и транспортера глюкозы 2 в абсорбции цианидин-3-O-β-глюкозида в клетках Caco-2. Питательные вещества. 2014;6:4165–4177. дои: 10.3390/nu6104165. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Активность переносчика глюкозы у млекопитающих зависит от анионных и конических фосфолипидов. Дж. Биол. хим. 2016; 291:17271–17282. doi: 10.1074/jbc.M116.730168. [Статья PMC бесплатно] [PubMed] [CrossRef] [Google Scholar]
54. De C., Palacio J.R., Martinez P., Morros A. Biochimica et Biophysica Acta Влияние окислительного стресса на текучесть плазматической мембраны THP-1, индуцированной макрофаги. BBA-Биомембр. 2013; 1828: 357–364. [PubMed] [Академия Google]
55. Phan H.T.T., Yoda T., Chahal B., Morita M., Takagi M., Vestergaard C. Biochimica et Biophysica Acta Структурно-зависимые взаимодействия полифенолов с биомиметической мембранной системой. BBA-Биомембр. 2014; 1838: 2670–2677. doi: 10.1016/j.bbamem.2014.07.001. [PubMed] [CrossRef] [Google Scholar]
BBA-Биомембр. 2014; 1838: 2670–2677. doi: 10.1016/j.bbamem.2014.07.001. [PubMed] [CrossRef] [Google Scholar]
56. Tsuchiya H. Мембранные взаимодействия фитохимических веществ как их молекулярный механизм, применимый к обнаружению выводов лекарств из растений. Молекулы. 2015;20:18923–18966. дои: 10.3390/молекулы201018923. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
При использовании местных растений в ландшафте выбор растений имеет значение
Местные цветы Keystone, такие как ароматические астры, поддерживают до 115 видов гусениц. Фото: Ферн Кэмпбелл. Ферн Кэмпбелл
Главный садовник Пьемонта
Во время Месяца местных растений в апреле жители Вирджинии и всей страны празднуют, сажая местные деревья, кустарники, многолетники, виноградные лозы и травы, чтобы создать среду обитания, которая позволит нашим птицам, опылителям , и дикая природа процветать. Включение местных растений в наши основные проекты садов необходимо, чтобы обратить вспять сокращение популяций наших птиц и насекомых и восстановить баланс в наших экосистемах.
Местные растения являются аборигенными для нашего региона и эволюционировали здесь на протяжении веков вместе с нашими местными насекомыми, птицами и другими животными. Вместе они делают регион Вирджиния-Пьемонт уникальным, обеспечивая диких животных пищей, укрытием и местами для выращивания детенышей, тем самым поддерживая биоразнообразие и формируя основу пищевой сети и здоровых экосистем.
Ставки высоки. Согласно исследованию, опубликованному в 2019 году журналом Science, общая популяция гнездящихся птиц в США и Канаде сократилась почти на 30 процентов с 19 века.70. Количество летающих насекомых, таких как мотыльки, сократилось более чем на 75 процентов с 1990-х годов. Насекомые опыляют 90 процентов наших цветковых растений, и без этих насекомых и других опылителей мы потеряем эти растения, и пищевая сеть рухнет.
Поскольку местные растения полностью адаптированы к нашему местному климату и почвам, они могут процветать в условиях, когда экзотические садовые растения испытывают трудности.
Оказывается, не всякое родное растение годится. Выбор растений имеет значение.
Исследования показали, что особый выбор местных растений, который делают садоводы, может оказать огромное влияние на разнообразие дикой природы в наших ландшафтах. Многолетние наблюдения и исследования энтомолога из Делавэрского университета Дугласа Таллами и его помощников показали, что определенные виды местных растений, которые он называет «краеугольными растениями», играют важную роль в поддержании здоровой пищевой сети, особенно для насекомых и животных, которые питаются на насекомых. Они также необходимы многим видам диких животных для завершения их жизненного цикла.
В каменной арке ключевой камень удерживает другие камни на месте. Убери его, и арка рухнет. В своей книге «Природа дубов » Таллами описывает, как краеугольные камни обеспечивают столь же важную поддержку для наших местных экосистем. «Двор без краеугольных растений, — пишет он, — будет далеко от обилия насекомых, необходимого для поддержания жизнеспособных пищевых сетей, даже если присутствуют десятки родов местных растений».
По данным Национальной федерации дикой природы, существует два типа ключевых растений:
1. Растения-хозяева, которыми питаются молодые гусеницы примерно 90 процентов бабочек, мотыльков и прыгунов (Lepidoptera) . Их листва служит пищей для богатых белком гусениц, которые, в свою очередь, являются важными компонентами рациона многих животных, в том числе птенцов большинства птиц.
2. Растения, которыми питаются пчелы-специалисты, питающиеся пыльцой только определенных видов. Установки Keystone для местных пчел кормят как специализированных, так и универсальных пчел.
Установки Keystone для местных пчел кормят как специализированных, так и универсальных пчел.
Исследование Таллами показало, что 90 процентов того, что едят гусеницы, создается только 14 процентами местных видов растений, и только пять процентов мощных растений получают 75 процентов пищи. «Возьмите краеугольное дерево, такое как дуб. Этот дуб могут съесть более 500 гусениц», — объясняет Таллами. «Это позволяет создать более сложную и стабильную пищевую сеть. Эти краеугольные камни или растения-электростанции, поддерживающие гусениц, выполняют большую часть работы, и без них пищевая сеть обречена».
В Среднеатлантическом регионе насчитывается более 2000 родов местных растений, из которых команда Таллами отнесла 38 к категории ключевых растений. Местные дубы ( Quercus ), дикие вишни ( Prunus ), ивы ( Salix ) и березы ( Betula ) возглавляют его список родов деревьев. К наиболее мощным травянистым растениям относятся золотарники ( Solidago ), астры ( Symphyotrichum ), земляника ( Fragaria ) и многолетние подсолнухи ( Подсолнечник ).
Таллами убедительно доказывает, что домовладельцы должны включить в свой ландшафт основные виды растений, произрастающих в нашем регионе. Поддерживая местные популяции насекомых, эти растения создают более продуктивные ландшафты, которые помогают восстанавливать экосистемы и поддерживать пищевую сеть. Вы должны выбрать разнообразие местных ключевых растений, чтобы оказать наибольшее влияние, и выбрать растения, подходящие для вашей существующей почвы, влажности, солнечного света и других условий участка.
Узколистный подсолнух обеспечивает цветение в конце сезона для опылителей. Фото: Билл Саблетт. Вы можете узнать больше о местных растениях в нашем регионе, заглянув во вкладку «Ресурсы для садоводства» на веб-сайте Мастеров-садоводов Пьемонта, piedmontmastergardeners.org. Среди прочего, вы найдете недавно обновленный список местных растений и приемлемых культурных сортов для региона Северного Пьемонта, а также список местных питомников и садовых центров, в которых продаются местные растения — розничных партнеров Plant Northern Piedmont Natives. Находясь там, вы можете загрузить такие ресурсы, как «Каждому саду нужны ключевые растения», в которых выделяются ключевые растения, произрастающие в нашем регионе, и серию списков растений для конкретных мест, разработанных PMG, с такими названиями, как «Крутые растения для горячих мест», «Прохладные растения для сухой тени», «Ковер для вашего сада» (почвопокровные) и «Местные растения для птиц».
Находясь там, вы можете загрузить такие ресурсы, как «Каждому саду нужны ключевые растения», в которых выделяются ключевые растения, произрастающие в нашем регионе, и серию списков растений для конкретных мест, разработанных PMG, с такими названиями, как «Крутые растения для горячих мест», «Прохладные растения для сухой тени», «Ковер для вашего сада» (почвопокровные) и «Местные растения для птиц».
Другие полезные онлайн-ресурсы включают поисковик растений Национальной федерации дикой природы (www.nwf.org/nativeplantfinder/), который предлагает справочник по местным растениям по почтовому индексу. А чтобы услышать мнение самого профессора Таллами, поищите его презентацию на You Tube «Местные краеугольные камни для дикой природы».
Опять же, выбор растения имеет значение. Вместо того, чтобы выбирать растение, которое просто приятно для человеческого глаза, выбирайте растения, которые также поддерживают сложную сеть жизни. Здоровые экосистемы обеспечивают основные услуги, от которых мы все зависим.