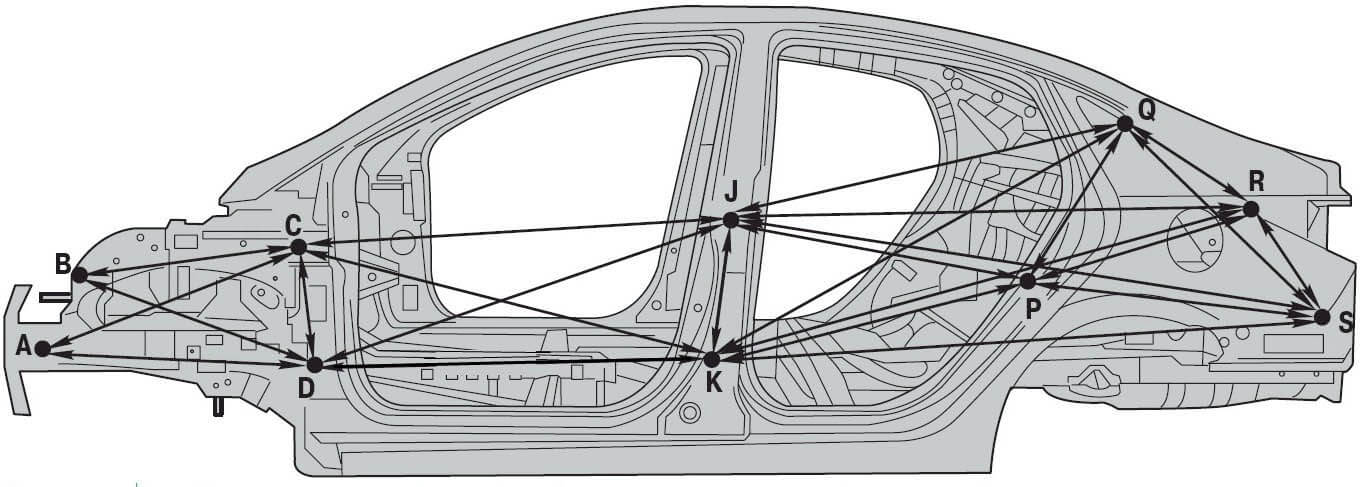
Контрольные геометрические размеры и точки кузова Nissan Juke / Ниссан Жук (джук)
Все приведенные на иллюстрациях размеры являются реальными (не проекционными).
При использовании приспособления для обмеров кузова, отрегулировать оба указателя на одинаковую длину. Убедиться в отсутствии посторонних люфтов в приспособлении.
• При использовании рулетки убедиться в отсутствии её растяжений, скручиваний или перегибов.
• Измерения должны проводиться от центра монтажных отверстий.
• Значения, отмеченные «*», указывают на то, что они соответствуют измерениям, проведенным с другой стороны автомобиля по симметричным меткам.
• Координаты мерных меток приведены в стандартной системе измерения по осям «X», «V» и «Z».
• Ось «Z» — воображаемая базовая линия (200 мм ниже базисной линии («0Z» на чертеже)).
1. Центр автомобиля. 2. Центр передней оси. 3. Воображаемая основная линия.
Центральные контрольные точки кузова
Центральные метки расположены на каждой части кузова для определения центра автомобиля.
| Точка | Участок | Метка |
| А | Передняя часть крыши | Насечка |
| В | Верхняя часть капота | Насечка |
| С | Верхняя часть перегородки | Насечка |
| D | Усиливающий элемент рычага переключения коробки передач | Отверстие Ø 16 мм |
| Е | Усиливающий элемент стояночного тормоза | Отверстие Ø 12 мм |
| F | Задняя панель | Выступ |
| G | Задняя часть крыши | Насечка |
Размеры передней части кузова, моторный отсек
Неуказанные размеры
(Версии с передним приводом)
| Точки измерения | Размер | Точки измерения | Размер | Точки измерения | Размер | Точки измерения | Размер |
| А-С | 729 (28.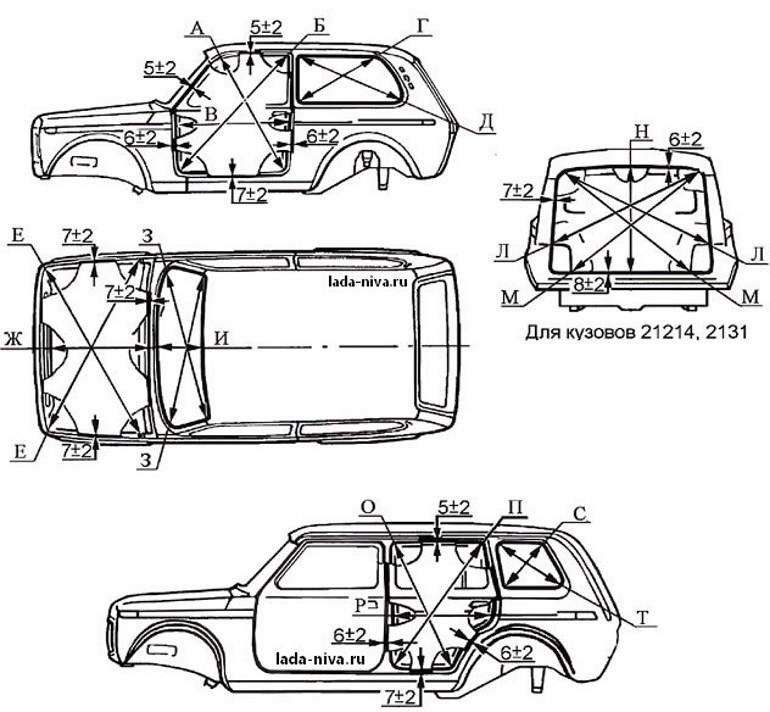 | A-J | 673 (26.50) | С-с | 1441 (56.73) | F-f | 1366 (53.78) |
| A-D | 729 (28.70)* | В-Е | 298 (11.73)* | D-d | 1429 (56.26) | G-h | 1192 (46.93)* |
| A-F | 798 (31.42)* | В-е | 1233 (48.54)* | Е-е | 1037 (40.83) | J-j | 965 (37.99) |
(Версии с полным приводом)
| Точки измерения | Размер | Точки измерения | Размер | Точки измерения | Размер | Точки измерения | Размер |
| А-С | 729 (28. 70)* 70)* | В-Е | 298 (11.73)* | D-d | 1429 (56.26) | G-h | 1192 (46.93)* |
| A-D | 729 (28.70)* | В-е | 1233 (48.54)* | Е-е | 1037 (40.83) | J-j | |
| A-F | 798 (31.42)* | С-с | 1441 (56.73) | F-f | 1366 (53.78) |
| Точка | Описание |
| А | Центральная метка кузова — насечка верхней части перегородки |
| В, b | Центр установочного отверстия навеса капота (14×10 мм) |
| С, с, F, f | Центр установочного отверстия переднего крыла (Ø 7 мм) |
| D,d | Центр установочного отверстия передней комбинированной фары головного освещения (Ø 7 мм) |
| Е, е | Центр установочного отверстия передней стойки 16×10 мм |
| G, g, Н, h | Центр установочного отверстия верхней опоры радиатора: G.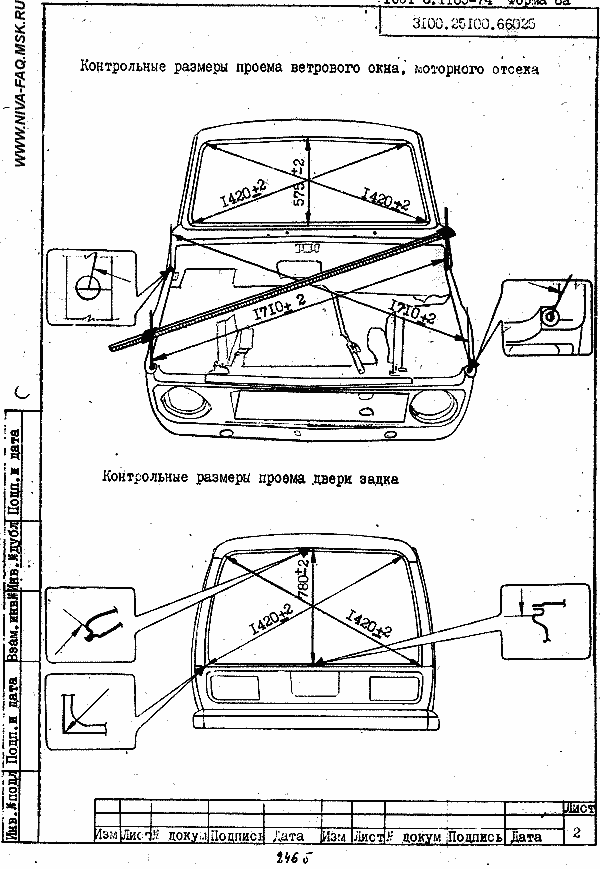 g: Ø 12 мм H, h: 14×12 мм g: Ø 12 мм H, h: 14×12 мм |
| J | Центр отверстия передней боковой балки (Ø 7 мм) |
| j | Центр отверстия монтажного кронштейна двигателя (Ø 11 мм) |
| К, k, М, m | Центр установочного отверстия стойки переднего бампера (Ø 15 мм) |
Размеры днища кузова
Версии с передним приводом (2WD)
| Точка | Координаты | Описание | ||
| Y | Z | |||
| А | 473.5(18.642) | -557.0 (-21.929) | 293.0 (11.535) | Отверстие Ø 11 мм |
| а | -491. 0 (-19.331) 0 (-19.331) | -557.0 (-21.929) | 301.9(11.886) | Отверстие Ø 11 мм |
| в | 472.4(18.598) | -455.0 (-17.913) | 328.9 (12.949) | Отверстие Ø 16 мм |
| b | -489.7 (-19.279) | -455.0 (-17.913) | 328.9 (12.949) | Отверстие Ø 18 мм |
| С, С | ±410.0 (±16.142) | 547.0 (21.535) | 190.2 (7.488) | Отверстие Ø 16 мм |
| D, d | ±410.0 (±16.142) | 815.0 (32.087) | 184.0 (7.244) | Отверстие Ø 14 мм |
| Ее | ±410.0 (±16.142) | 1173.0 (46.181) | 170.1 (6.697) | Отверстие 14×2 мм |
| F, f | ±633.0 (±24.921) | 2032.3 (80.012) | 318.0(12.520) | Отверстие Ø 16 мм |
| G,g | ±350.0 (±13.780) | 2253. 0 (88.701) 0 (88.701) | 390.4(15.370) | Отверстие Ø 11 мм |
| Н, h | ±482.3 (±18.988) | 2374.2 (93.472) | 348.3 (13.713) | Отверстие Ø 12 мм |
| J | 499.5 (19.665) | 2448.0 (96.378) | 378.0 (14.882) | Отверстие Ø 21 мм |
| i | -487.5 (-19.193) | 2448.0 (96.378) | 378.0 (14.882) | Отверстие Ø 21 мм |
| К | 498.0 (19.606) | 2743.0 (107.992) | 378.0 (14.882) | Отверстие 17×16 мм |
| k | -487.5 (-19.193) | 2743.0 (107.992) | 378.0 (14.882) | Отверстие 17×16 мм |
| M | 498.0 (19.606) | 2970.0(116.929) | 375.4(14.779) | Отверстие Ø 16 мм |
| m | -485.1 (-19.098) | 2965.8(116.764) | 375.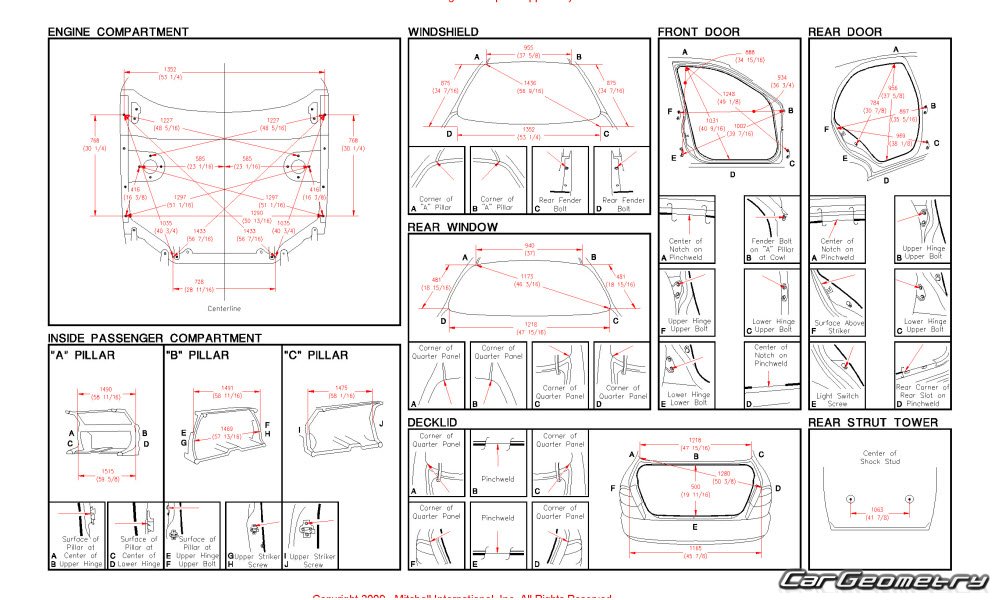 4 (14 779) 4 (14 779) | Отверстие Ø 16 мм |
| N, n | ±583.6 (±22.976) | 6.6 (0.261) | 868.2(34.181) | Отверстие Ø 98 мм |
| O,o | ±575.4 (±22.653) | 2569.2(101 149) | 734.4 (28.913) | Отверстие Ø 20 мм |
Версии с полным приводом (4WD)
| Точка | Координаты | Описание | ||
| X | Y | Z | ||
| А | 473.5 (18.642) | -557.0 (-21.929) | 293.0(11.535) | Отверстие Ø 11 мм |
| а | -491. 0 (-19.331) 0 (-19.331) | -557.0 (-21.929) | 301.9(11.886) | Отверстие Ø 11 мм |
| в | 472.4 (18.598) | -455.0 (-17.913) | 328.9 (12.949) | Отверстие Ø 18 мм |
| b | -489.7 (-19.279) | -455.0 (-17.913) | 328.9 (12.949) | Отверстие Ø 18 мм |
| С, с | ±410.0 (+16.142) | 547.0 (21.535) | 190.2 (7.488) | Отверстие Ø 16 мм |
| D, d | ±410.0 (±16.142) | 815.0 (32.037) | 184.0 (7.244) | Отверстие Ø 12 мм |
| Е е | ±410.0 (±16.142) | 1173.0 (46.181) | 170.1 (6.697) | Отверстие 14×2 мм |
| F, f | ±601.8 (±23.693) | 2016.4(79.386) | 315.0(12.402) | Отверстие Ø 15 мм |
| G,g | ±515.1 (±20.279) | 2318.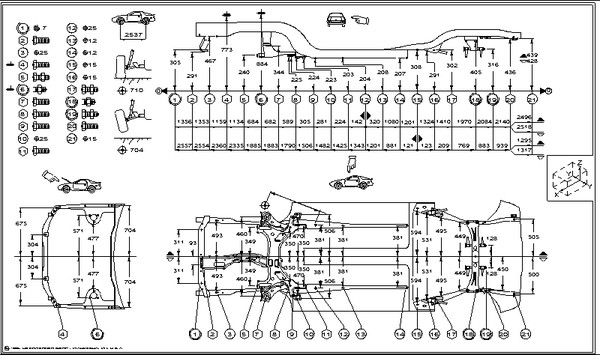 6 (91.283) 6 (91.283) | 357.2 (14.063) | Отверстие Ø 12 мм |
| Н, h | ± 330.0 (±12.992) | 2339.7 (92.114) | 419.0 (16.496) | Отверстие 33×30 мм |
| J, i | ±482.0 (±18.976) | 2648.0 (104.252) | 422.2 (16.622) | Отверстие 20×6 мм |
| К, k | ±300.0 (±11.811) | 2815.0(110.827) | 417.2(16.425) | Отверстие 18×6 мм |
| М, m | ±482.7 (±19.004) | 2910.5(114.586) | 421.4(16.591) | Отверстие Ø 23 мм |
| N, n | ±583.6 (±22.976) | 6.6 (0.261) | 868.2(34.181) | Отверстие Ø 98 мм |
| O, о | ±558.6 (±22.000) | 2568.4(101.118) | 734.4(28.913) | Отверстие Ø 20 мм |
Размеры проёмов дверей и лобового стекла
Неуказанные размеры
| Точки измерения | Размер | Точки измерения | Размер | Точки измерения | Размер | Точки измерения | Размер |
| А-Е | 724(28.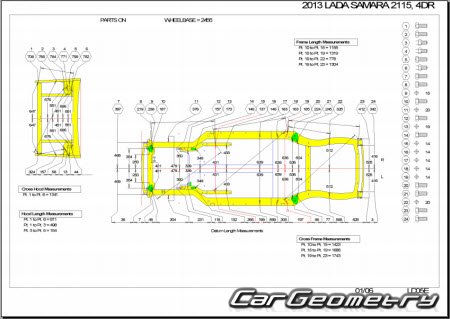 50)* 50)* | H-h | 1406 (55.35) | N-n | 1423 (56.02) | R-J | 1394 (54.88)* |
| В-Е | 1028 (40.47)* | H-j | 1829 (72.01)* | N-o | 1585 (62.40)* | R-K | 1072 (42.20)* |
| C-d | 1290 (50.79)* | H-k | 1663 (65.47)* | N-p | 1554(61 18)* | S-M | 1094 (43.07)* |
| С-е | 1423 (55.02)* | J-J | 1257 (49.49) | N-q | 1506 (59. 29)* 29)* | S-N | 786 (30.94)* |
| F-f | 1222 (48.11) | J-k | .1463 (57.60)* | O-o | 1326 (52.20) | S-0 | 1248 (49.13)* |
| F-h | 1569 (61 77)* | K-k | 1425 (56.10) | Oq | 1518 (59.76)* | S-P | 1126(44.33)* |
| F-j | 1386 (54.57)* | M-m | 1248 (49.13) | P-p | 1397 (55.00) | S-0 | 905 (35.63)* |
| F-k | 1519 (59.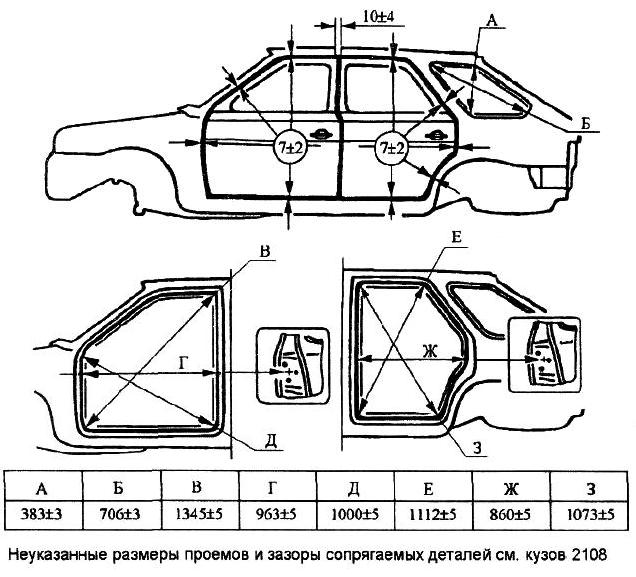 80)* 80)* | M-n | 1451 (5713)* | G-q | 1444 (56.85) | T-V | 1128 (44.41)* |
| G-g | 1359 (53.50) | M-o | 1441 (56.73)* | R-F | 1104 (43.46)* | T-W | 1105(43.50)* |
| G-j | 1630 (64.17)* | M-p | 1486 (58.50)* | R-G | 885 (34.84)* | U-V | 1204 (47.40)* |
| G-k | 1616 (63.62)* | M-q | 1578 (62.13)* | R-H | 720 (28. 35)* 35)* | U-W | 1091 (42.95)* |
| Точка | Описание |
| А | Центральная метка кузова — насечка верхней части перегородки |
| В | Центральная метка кузова — выпуклость на краю крыши |
| С, с | Соединительный шип снаружи боковины кузова |
| D, d, G, g | Соединительный шип внутри передней стойки |
| Е, е | Центр установочного отверстия навеса капота (Ø 12 мм) |
| F, f, Н, h | Выпуклость передней стойки |
| J, j, К, k, М, т, N, n | Выпуклость центральной стойки |
| О, о,Q, q | Выпуклость заднего крыла |
| Р, p | Соединительный шип снаружи задней колесной арки |
| R | Центральная метка кузова — центр отверстия усиливающего элемента рычага переключения коробки передач (Ø 16 мм) |
| S | Центральная метка кузова — центр отверстия усиливающего элемента рычага стояночного тормоза (Ø 12 мм) |
| Т, t, U, u V, V, W, w | Центр установочного отверстия дверного навеса: T,t, U, u, W, w: Ø 12 мм V, v: Ø 9 мм |
Размеры проёма багажного отделения, задняя часть кузова
| Точка | Описание |
| А | Центральная метка кузова — выпуклость на краю крыши |
| В, b | Соединительный шип продолжения заднего крыла |
| С, с D, d | Соединительный шип основания заднего комбинированного фонаря |
| Е | Центральная метка кузова — выпуклость задней поперечины |
Контрольные размеры геометрии кузова — Кузовщина
#1 Дизель 2.
 0
0Отправлено 15 Сентябрь 2018 — 23:35
Здравствуйте!сложилась такая ситуация,при разборе передней части авто,увидел что ланжероны по отнашению к передней рамке,ушли в лево, примерно на два сантима.когда покупал,видно ни чего небыло.у меня вопрос:где взять размеры кузова.что бы вытянуть лонжероны?
- Наверх
#2 stilnyyp
Отправлено 08 Октябрь 2018 — 22:27
Хорошему кузовщику размеры не нужны.
Dodge Journey 2010 SXT 3.5 AWD 7seats один хозяин
- Наверх
#3 TRS-motors
Отправлено 09 Октябрь 2018 — 10:53
stilnyyp, не порите чушь. Ваше утверждение сродни «хорошему мотористу динамометрический ключ не нужен». Чтобы не было п*здежу, делай всё по чертежу – основа основ.
Ваше утверждение сродни «хорошему мотористу динамометрический ключ не нужен». Чтобы не было п*здежу, делай всё по чертежу – основа основ.
Дизель 2.0, нужные файлы во вложении. Удачи.
Прикрепленные файлы
- Frame Dimensions.pdf 350,21К 221 Количество загрузок:
- Opening Dimensions.pdf 630,27К 197 Количество загрузок:
- rehmzy, Прохожий, Андрей Гостенков и еще 1 это нравится
- Наверх
#4 Дизель 2.0
Отправлено 19 Октябрь 2018 — 01:47
Спасибо огромное!!!очень помогли
- Наверх
#5 Konsstantin
Отправлено 06 Июнь 2020 — 09:23
Подскажите, а эти размеры подходят под crossroad 2,4 2017? Меня интересует передок. Был удар в переднее правое колесо, повело ланжероны и немного стакан.
Был удар в переднее правое колесо, повело ланжероны и немного стакан.
Сообщение отредактировал Konsstantin: 06 Июнь 2020 — 09:30
- Наверх
#6 fedain
Отправлено 20 Апрель 2021 — 12:15
Вот и я получил авто, а лонжероны ушли ((
Подскажите пожалуйста, 2018 se 2.4, USA подходят указанные выше?
- Наверх
#7 URRI 39
Отправлено 20 Апрель 2021 — 12:42
Конечно подходят, кузов то один.
- Наверх
#8 fedain
Отправлено 20 Апрель 2021 — 12:49
Спасибо !
Сообщение отредактировал MoparPlus: 20 Апрель 2021 — 15:03
Оверквотинг.
- Наверх
Большой или быстрый: две стратегии контроля над размерами тела в процессе развития | BMC Biology
- Комментарий
- Открытый доступ
- Опубликовано:
- Х. Фредерик Нейхаут 1
Биология BMC том 13 , номер статьи: 57 (2015) Процитировать эту статью
3164 доступа
17 цитирований
1 Альтметрический
Сведения о показателях
Abstract
Размеры тела взрослых особей контролируются механизмами, которые останавливают рост при достижении характерных для вида размеров. Механизмы, с помощью которых ощущается размер и с помощью которых эта информация передается в систему регуляции роста, начинают понимать у некоторых видов насекомых. Были обнаружены две довольно разные стратегии контроля; один благоприятствует крупным размерам тела, а другой способствует быстрому развитию.
Механизмы, с помощью которых ощущается размер и с помощью которых эта информация передается в систему регуляции роста, начинают понимать у некоторых видов насекомых. Были обнаружены две довольно разные стратегии контроля; один благоприятствует крупным размерам тела, а другой способствует быстрому развитию.
Комментарий
Возраст и размер в зрелом возрасте, возможно, являются двумя наиболее важными чертами жизненного цикла животных. Варьирование обоих признаков оказывает серьезное влияние на приспособленность: возраст зрелости влияет на время поколения, а размер зрелости оказывает сильное влияние на репродуктивную способность. Соответственно, возраст и размер во взрослом возрасте интенсивно изучались с эволюционной и экологической точек зрения [1, 2]. Тем не менее, несмотря на их очевидную важность, генетические, онтогенетические и физиологические механизмы, которые контролируют возраст и размер в зрелом возрасте, по большей части неизвестны. Питание и гормоны играют очевидную и хорошо зарекомендовавшую себя роль в росте, но естественные механизмы, вызывающие прекращение роста, когда животное достигает видоспецифического размера, остаются одной из величайших загадок биологии.
В недавней статье Hatem et al. [3] пролили новый свет на механизмы развития и физиологические механизмы, которые регулируют рост и размер тела, и в то же время раскрыли причину загадочного и неприятного различия в механизмах между двумя видами, Manduca sexta и Drosophila melanogaster . , в которых управление размером изучено лучше всего.
Для роста до видового размера требуется механизм, который может контролировать размер, и механизм реагирования, который останавливает рост и срабатывает при достижении определенного размера. Такие механизмы контроля размера и реакции не известны ни у одного млекопитающего, но теперь они начинают хорошо пониматься у некоторых насекомых, в частности у M. sexta , бабочка, и D. melanogaster , муха. Непосредственный триггер прекращения роста насекомых хорошо известен, а именно импульс стероидного гормона экдизона, возникающий в конце личиночной жизни. Этот пульс заставляет животное перестать питаться и начать метаморфическую линьку. Насекомые не растут во взрослом состоянии, поэтому размер личинки во время этого импульса экдизона полностью определяет размер взрослого насекомого.
Насекомые не растут во взрослом состоянии, поэтому размер личинки во время этого импульса экдизона полностью определяет размер взрослого насекомого.
Откуда вы знаете, насколько вы велики?
Контроль времени этого импульса экдизона был впервые исследован в Manduca [4, 5]. Примерно в середине пятого и последнего личиночного возраста личинки Manduca преодолевают так называемый критический вес. Это размер, при котором запускаются физиологические процессы, приводящие к секреции экдизона. В течение этого возраста секреция экдизона подавляется ювенильным гормоном (ЮГ), а при критическом весе секреция ЮГ прекращается, а экспрессия фермента ЮГ-эстеразы (основного катаболического фермента ЮГ) усиливается. Подавление секреции экдизона снимается в течение 24–60 часов (в зависимости от генетического фона). Таким образом, окончательный размер личинки задается величиной критической массы и продолжительностью периода элиминации ЮГ, называемого терминальной фазой роста (ТФР).
Механизм контроля размера, который определяет критический вес, был довольно неожиданным и включает ограничение кислорода нерастущей системой трахеи, которая становится неспособной справляться с постоянно растущими потребностями растущего организма в кислороде. Размер, при котором кислород становится предельным, соответствует критическому весу [6]. Выращивание личинок Manduca в условиях гипоксии снижает их критическую массу, а также уменьшает размер, при котором секретируется экдизон и прекращается рост. Аналогичный механизм работает в Drosophila , где личинки, выращенные в условиях гипоксии, также дают миниатюрных взрослых мух [7], предположительно из-за преждевременного запуска метаморфической линьки. У обоих видов первым шагом каскада, регулирующего размер тела, является ограничение кислорода. В Manduca ограничение кислорода определяет критический вес и вызывает снижение JH. У Drosophila JH не играет роли в регуляции секреции экдизона.
Одно из самых ярких отличий между Manduca и Drosophila заключается в том, что Drosophila , по-видимому, практически не имеет ЮГ на третьей и последней личиночной стадии. А поскольку у Manduca JH играет ключевую роль в регуляции размеров тела, естественно было предположить, что у Drosophila контроль размеров должен быть совсем другим. Эта идея была подкреплена открытием того, что вмешательство в передачу сигналов инсулина может оказывать сильное влияние на размер взрослого тела у Drosophila . Подавление продукции инсулина или инактивация рецептора инсулина или субстрата рецептора приводили к появлению миниатюрных мух [8], тогда как избыточная экспрессия инсулина приводила к появлению гигантских мух [9].].
А поскольку у Manduca JH играет ключевую роль в регуляции размеров тела, естественно было предположить, что у Drosophila контроль размеров должен быть совсем другим. Эта идея была подкреплена открытием того, что вмешательство в передачу сигналов инсулина может оказывать сильное влияние на размер взрослого тела у Drosophila . Подавление продукции инсулина или инактивация рецептора инсулина или субстрата рецептора приводили к появлению миниатюрных мух [8], тогда как избыточная экспрессия инсулина приводила к появлению гигантских мух [9].].
Еще одно интересное различие между Manduca и Drosophila заключается в их реакции на голодание после достижения критического веса. Когда личинки Manduca голодают после того, как они преодолели критический вес, их время секреции экдизона и окукливания идентично времени личинок, которые продолжают нормально питаться [5]. Напротив, когда личинок дрозофилы голодают, их время до окукливания ускоряется [10]. Эта преждевременная метаморфоза была названа механизмом спасения [11]. Это обычная адаптация у животных, живущих в мимолетных местах обитания, где невозможно найти новый источник пищи после того, как текущий иссякнет, хотя механизм экстренной реакции до сих пор неизвестен.
Эта преждевременная метаморфоза была названа механизмом спасения [11]. Это обычная адаптация у животных, живущих в мимолетных местах обитания, где невозможно найти новый источник пищи после того, как текущий иссякнет, хотя механизм экстренной реакции до сих пор неизвестен.
Все виды не одинаковы
Из-за этих различий в эндокринной биологии двух видов работа по регуляции размера у Manduca была сосредоточена в основном на физиологии критического веса и роли ЮГ в контроле ecdysone, в то время как у Drosophila большая часть работы была сосредоточена на роли передачи сигналов инсулина и относительных ролях членов сети передачи сигналов инсулина, таких как FOXO и TOR [12].
Хотя многие аспекты морфологии и физиологии Drosophila являются высокоразвитыми по сравнению с другими насекомыми, казалось трудным понять, почему Drosophila развила такие радикально отличающиеся механизмы контроля над размером тела, чем те, что были обнаружены в Manduca . Эта головоломка могла бы быть решена, если бы был только способ сделать Drosophila более похожим на Manduca или Manduca более похожим на Drosophila .
Эта головоломка могла бы быть решена, если бы был только способ сделать Drosophila более похожим на Manduca или Manduca более похожим на Drosophila .
Последнее было выполнено в статье Hatem et al. [3]. Оказывается, существует генетический штамм Manduca , черный штамм, который, помимо наличия черных личинок и гораздо меньшего размера взрослого тела, имеет сниженный уровень ЮГ на личиночной стадии [13]. В этом последнем отношении он напоминает Drosophila .
What Hatem et al. [3] заключалась в том, чтобы кормить личинок дикого типа и черных личинок диетой, содержащей рапамицин, который ингибирует передачу сигналов TOR, один из конечных этапов инсулинового пути. Они обнаружили, что, хотя рост обоих штаммов замедлился, как и можно было ожидать, штамм дикого типа в конечном итоге вырос до нормального размера, тогда как черный штамм стал намного больше, чем обычно. Интересно, что критический вес не изменился ни у одного из штаммов при введении рапамицина. Следовательно, увеличение размеров черных личинок должно было происходить за счет удлинения TGP.
Следовательно, увеличение размеров черных личинок должно было происходить за счет удлинения TGP.
TOR контролирует не только восприятие аминокислот, синтез белка и рост клеток в тканях, но и биосинтез экдизона в переднегрудных железах (рис. 1). Задержка линьки предположительно была связана с тем, что вмешательство в TOR также мешало и задерживало секрецию экдизона. Хатем и др. [3] предполагают, что рапамицин не оказывал такого эффекта на штамм дикого типа, потому что его эффект подавляется высокими уровнями ЮГ в этом штамме.
Рис. 1 Инсулиновая сигнальная сеть, которая связывает питание с ростом и размером. Питание в виде аминокислот стимулирует синтез инсулина. Активация рецептора инсулина и субстрата рецептора (INR/INS) активирует PKC и PKB/Akt посредством многоэтапного пути. ПКС стимулирует поглощение аминокислот, которые активируют TOR. Инсулин также активирует TOR через PKB/Akt. TOR ингибируется FOXO, который ингибируется PKB/Akt. TOR стимулирует синтез белка через инициатор транскрипции EIF4E и вызывает рост клеток в тканях. В переднегрудных железах TOR стимулирует синтез экдизона. Экдизон ингибируется FOXO при связывании с Ultraspiracle (Usp) [16]. Ингибирование TOR рапамицином ингибирует рост и синтез экдизона. Экдизон также ингибируется ювенильным гормоном, который начинает распадаться при критическом весе из-за ограничения кислорода. Таким образом, окончательный контроль в этой сети обеспечивается двумя факторами окружающей среды: питанием и ограничением кислорода 9.0023
TOR стимулирует синтез белка через инициатор транскрипции EIF4E и вызывает рост клеток в тканях. В переднегрудных железах TOR стимулирует синтез экдизона. Экдизон ингибируется FOXO при связывании с Ultraspiracle (Usp) [16]. Ингибирование TOR рапамицином ингибирует рост и синтез экдизона. Экдизон также ингибируется ювенильным гормоном, который начинает распадаться при критическом весе из-за ограничения кислорода. Таким образом, окончательный контроль в этой сети обеспечивается двумя факторами окружающей среды: питанием и ограничением кислорода 9.0023
Изображение в полный размер
Еще одно интересное свойство черного штамма заключается в том, что когда личинки, которых кормили рапамицином, голодают, время их метаморфоза ускоряется, точно так же, как это наблюдается в реакции катапультирования у Drosophila , но которая не происходит в штамме дикого типа Manduca . Таким образом, в некоторых отношениях физиология развития черного штамма больше похожа на таковую у Drosophila , чем у дикого типа Manduca 9. 0020 .
0020 .
Две разные стратегии
Hatem et al. [3] предполагают, что должно быть два уровня контроля над экдизоном. Первый в первую очередь чувствителен к питанию и опосредован передачей сигналов инсулина/TOR и запускается при минимальном жизнеспособном весе (MVW), который запускает таймер задержки (механизм которого до сих пор неизвестен), что приводит к секреции экдизона [14]. Это механизм, который работает у дрозофилы . Второе происходит в Manduca , где JH преобладает над механизмом, опосредованным инсулином/TOR. Manduca также имеет MVW, который находится намного ниже критического веса [15] и работает только в стрессовых условиях [14]. Обычно снижение ЮГ начинается при критическом весе, и запускается другой таймер задержки (в данном случае время, необходимое для устранения ЮГ), что в конечном итоге приводит к секреции экдизона (рис. 2).
Рис. 2 Альтернативный механизм управления размером корпуса. Сигмоидальные кривые роста с двумя разными механизмами остановки роста. а В присутствии ювенильного гормона (ЮГ), как и в Manduca , секреция экдизона не может происходить до тех пор, пока ЮГ не исчезнет. Снижение ЮГ начинается, когда личинки преодолевают критическую массу (КВ). b В отсутствие JH, как и у Drosophila , секреция экдизона следует за достижением минимального жизнеспособного веса (MVW) после задержки, называемой таймером линьки, механизм которой еще не изучен. Это приводит к более раннему прекращению роста и меньшему размеру тела, чем при ЮГ, подавляющем механизм МВВ 9.0023
а В присутствии ювенильного гормона (ЮГ), как и в Manduca , секреция экдизона не может происходить до тех пор, пока ЮГ не исчезнет. Снижение ЮГ начинается, когда личинки преодолевают критическую массу (КВ). b В отсутствие JH, как и у Drosophila , секреция экдизона следует за достижением минимального жизнеспособного веса (MVW) после задержки, называемой таймером линьки, механизм которой еще не изучен. Это приводит к более раннему прекращению роста и меньшему размеру тела, чем при ЮГ, подавляющем механизм МВВ 9.0023
Изображение полного размера
Интересно предположить, какая из этих систем управления является примитивной. Поскольку чувствительность к JH и критический вес возникают позже в развитии, чем контроль, чувствительный к питанию, и MVW, похоже, что они могут иметь более позднее эволюционное происхождение. В качестве альтернативы, это могло быть так, что дрозофила , движимая адаптивным значением способности выручать из ухудшающейся среды, отказалась от контроля со стороны JH.
Хатем и др. [3] предполагают, что механизм, контролируемый ЮГ, способствует большому размеру тела, но за счет более длительного развития, тогда как устранение подавления секреции экдизона ЮГ способствует быстрому времени развития. Таким образом, различия между механизмами управления в Manduca и Drosophila , скорее всего, являются адаптациями для очень разных жизненных историй.
Какая из этих стратегий, вероятно, является конечной причиной различия, еще предстоит выяснить. Тем временем Хатем и соавт. [3] сделали значительный шаг в разрешении загадочных различий между Manduca и Drosophila , и эта работа закладывает основу для новых экспериментальных подходов к более полному раскрытию деталей физиологии развития регуляции размеров тела.
Ссылки
Stearns SC. Эволюция историй жизни. Оксфорд: Издательство Оксфордского университета; 1992.
Google Scholar
 «>
«>Рофф Д.А. Эволюция историй жизни. Нью-Йорк: Чепмен и Холл; 1992.
Google Scholar
Hatem N, Wang Z, Nave K, Koyama T, Suzuki Y. Роль ювенильного гормона и передачи сигналов инсулина/TOR в росте Manduca sexta. БМС Биол. 2015;13:44.
Артикул ПабМед Центральный пабмед Google Scholar
Нейхаут ХФ. Роль экдизона в окукливании Manduca sexta. Дж. Физиология насекомых. 1976; 22: 453–63.
Артикул КАС Google Scholar
Нейхаут Х.Ф., Уильямс CM. Контроль линьки и метаморфоза у табачного рогатого червя Manduca sexta (L.): прекращение секреции ювенильного гормона как триггер для окукливания. J Эксперт Биол. 1974;61:493–501.
КАС пабмед Google Scholar
 «>
«>Callier V, Nijhout HF. Контроль размера тела с помощью снабжения кислородом выявляет зависящие от размера и независимые от размера механизмы линьки и метаморфоза. Proc Natl Acad Sci U S A. 2011;108:14664–9.
Артикул КАС ПабМед Центральный пабмед Google Scholar
Генрих Э.К., Фарзин М., Клок С.Дж., Харрисон Дж.Ф. Влияние стадии развития на чувствительность размеров клеток и тела к гипоксии у Drosophila melanogaster. J Эксперт Биол. 2011;214:1419–7.
Артикул ПабМед Центральный пабмед Google Scholar
Бёни Р., Рисго-Эсковар Дж., Олдхэм С., Броджиоло В., Стокер Х., Андрусс Б.Ф. и др. Автономный контроль размера клеток и органов с помощью CHICO, гомолога дрозофилы IRS1-4 позвоночных. Клетка. 1999; 97: 865–75.
Артикул пабмед Google Scholar
 «>
«>Броджиоло В., Стокер Х., Икея Т., Ринтелен Ф., Фернандес Р., Хафен Э. Эволюционно законсервированная функция рецептора инсулина дрозофилы и инсулиноподобных пептидов в контроле роста. Карр Биол. 2001; 11: 213–21.
Артикул КАС пабмед Google Scholar
Мирт С., Трумэн Дж.В., Риддифорд Л.М. Роль переднегрудной железы в определении критического веса для метаморфоза у Drosophila melanogaster. Карр Биол. 2005; 15: 1796–807.
Артикул КАС пабмед Google Scholar
Тоблер А, Нейхаут Х.Ф. Переключение в контроле роста имагинальных дисков крыльев Manduca sexta. ПЛОС Один. 2010;5:e10723.
Артикул ПабМед Центральный пабмед Google Scholar
Нейхаут Х.Ф., Риддифорд Л.М., Мирт С., Шинглтон А.В., Сузуки Ю., Каллиер В. Контроль размера насекомых в процессе развития.
 Wiley Interdiscip Rev Dev Biol. 2014;3:113–34.
Wiley Interdiscip Rev Dev Biol. 2014;3:113–34.Артикул ПабМед Центральный пабмед Google Scholar
Сафранек Л., Риддифорд Л.М. Биология черного личиночного мутанта табачного рогового червя. Мандука секста. Дж. Физиология насекомых. 1975; 21: 1931–1938.
Артикул Google Scholar
Судзуки Ю., Кояма Т., Хирума К., Риддифорд Л.М., Трумэн Дж.В. Таймер линьки участвует в метаморфической линьке личинок Manduca sexta. Proc Natl Acad Sci U S A. 2013;110:12518–25.
Артикул КАС ПабМед Центральный пабмед Google Scholar
Callier V, Nijhout HF. Определение размера тела у насекомых: обзор и синтез механизмов, зависящих от размера и мозга, и независимых механизмов. Биол Рев. 2013; 88:944–54.
Артикул пабмед Google Scholar
 «>
«>Кояма Т., Родригес М.А., Атанасиадис А., Шинглтон А.В., Мирт К.К. Пищевой контроль размера тела посредством биосинтеза экдизона, опосредованного FoxO-Ultraspiracle. электронная жизнь. 2014;3:e03091.
Артикул ПабМед Центральный Google Scholar
Ссылки на скачивание
Информация об авторе
Авторы и организации
Отдел биологии, Университет Дьюка, Дарем, Северная Каролина, 27708, США
H. Frederik Nijhout
Авторы
- H. Frederik Nijhout
. PubMed Google Scholar
Автор, ответственный за корреспонденцию
Х. Фредерик Нейхут.
Права и разрешения
Открытый доступ Эта статья распространяется в соответствии с условиями международной лицензии Creative Commons Attribution 4.0 (http://creativecommons.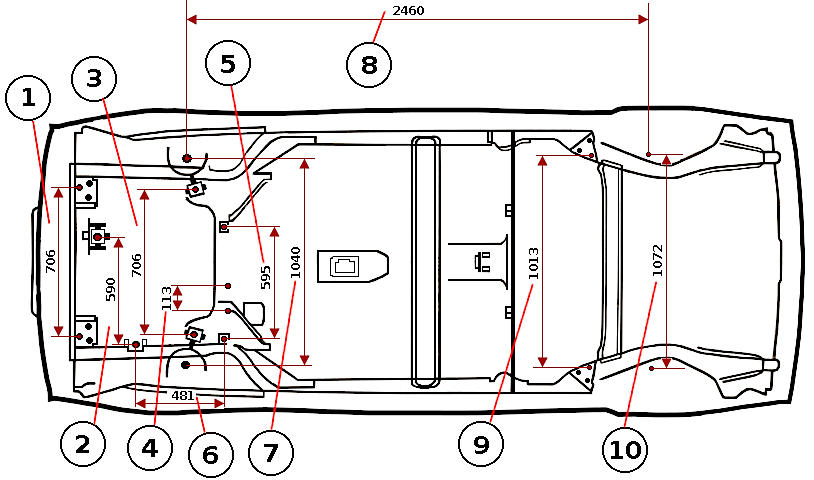 org/licenses/by/4.0/), которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии вы должным образом указываете автора (авторов) и источник, предоставляете ссылку на лицензию Creative Commons и указываете, были ли внесены изменения. Отказ от права Creative Commons на общественное достояние (http://creativecommons.org/publicdomain/zero/1.0/) применяется к данным, представленным в этой статье, если не указано иное.
org/licenses/by/4.0/), которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии вы должным образом указываете автора (авторов) и источник, предоставляете ссылку на лицензию Creative Commons и указываете, были ли внесены изменения. Отказ от права Creative Commons на общественное достояние (http://creativecommons.org/publicdomain/zero/1.0/) применяется к данным, представленным в этой статье, если не указано иное.
Перепечатка и разрешения
Об этой статье
Размер шрифта CSS
❮ Назад Далее ❯
Размер шрифта
Свойство font-size устанавливает размер текста.
Возможность управлять размером текста очень важна в веб-дизайне. Однако вы не следует использовать настройки размера шрифта, чтобы абзацы выглядели как заголовки, или заголовки выглядят как абзацы.
Всегда используйте правильные теги HTML, такие как
—
для заголовков и
для абзацы.
Значение размера шрифта может быть
абсолютный или относительный размер.
Абсолютный размер:
- Устанавливает для текста заданный размер
- Не позволяет пользователю изменять размер текста во всех браузерах (плохо по причинам доступности)
- Абсолютный размер полезен, когда известен физический размер вывода
Относительный размер:
- Устанавливает размер относительно окружающих элементов
- Позволяет пользователю изменять размер текста в браузерах
Примечание: Если не указать размер шрифта, размер по умолчанию для обычного текста, например абзацев, составляет 16 пикселей (16 пикселей = 1 см).
Установка размера шрифта в пикселях
Установка размера текста в пикселях дает вам полный контроль над размером текста:
Пример
h2 {
размер шрифта: 40 пикселей;
}
h3 {
Размер шрифта: 30 пикселей;
}
p {
размер шрифта: 14 пикселей;
}
Попробуйте сами »
Совет: Если вы используете пиксели, вы все равно можете использовать инструмент масштабирования для изменения размера всей страницы.
Установка размера шрифта с помощью Em
Чтобы позволить пользователям изменять размер текста (в меню браузера), многие разработчики используют em вместо пикселей.
1em равен текущему размеру шрифта. Размер текста по умолчанию в браузерах 16 пикселей. Таким образом, размер 1em по умолчанию составляет 16 пикселей.
Размер можно рассчитать от пикселей до em по следующей формуле: пикселей /16= em
Пример
h2 {
размер шрифта: 2,5 em; /* 40px/16=2.5em */
}
h3 {
размер шрифта: 1.875em; /* 30px/16=1.875em */
}
p {
font-size: 0.875em; /* 14px/16=0.875em */
}
Попробуйте сами »
В приведенном выше примере размер текста в em такой же, как и в предыдущем примере в пикселях. Однако с размером em можно настроить размер текста. во всех браузерах.
К сожалению, со старыми версиями все еще есть проблема
из Internet Explorer.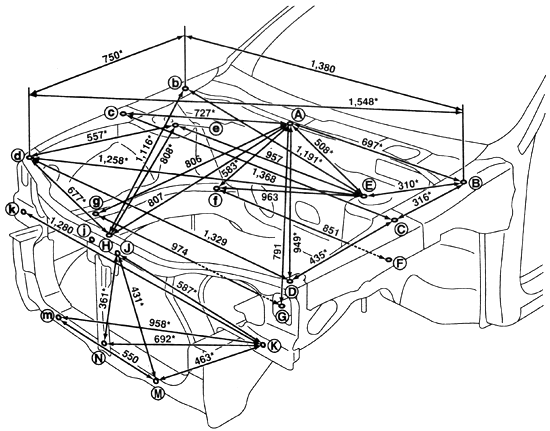 Текст становится больше, чем должен
при увеличении и меньше, чем должно, при уменьшении.
Текст становится больше, чем должен
при увеличении и меньше, чем должно, при уменьшении.
Используйте комбинацию Percent и Em
Решение, которое работает во всех браузерах, заключается в установке размера шрифта по умолчанию в процент для элемента
:Пример
body {
размер шрифта: 100%;
}
h2 {
размер шрифта: 2,5 em;
}
h3 {
размер шрифта: 1,875 em;
}
p {
размер шрифта: 0,875 em;
}
Попробуйте сами »
Теперь наш код отлично работает! Он показывает тот же размер текста в все браузеры, и позволяет всем браузерам масштабировать или изменять размер текста!
Размер адаптивного шрифта
Размер текста можно задать в единицах vw , что означает «ширину окна просмотра».
Таким образом, размер текста будет соответствовать размеру окна браузера:
Измените размер окна браузера, чтобы увидеть, как масштабируется размер шрифта.
Пример
Hello World
Попробуйте сами »
Viewport — размер окна браузера. 1vw = 1% ширины области просмотра. Если окно просмотра имеет ширину 50 см, 1vw составляет 0,5 см.
❮ Предыдущий Следующий ❯
ВЫБОР ЦВЕТА
Лучшие учебники
Учебное пособие по HTMLУчебное пособие по CSS
Учебное пособие по JavaScript
Учебное пособие
Учебное пособие по SQL
Учебное пособие по Python
Учебное пособие по W3.CSS
Учебное пособие по Bootstrap
Учебное пособие по PHP
Учебное пособие по Java
Учебное пособие по C++
Учебное пособие по jQuery
Справочник по HTML
Справочник по CSS
Справочник по JavaScript
Справочник по SQL
Справочник по Python
Справочник по W3.CSS
Справочник по Bootstrap
Справочник по PHP
Цвета HTML
Справочник по Java
Справочник по Angular
Справочник по jQuery
Основные примеры
Примеры HTMLПримеры CSS
Примеры JavaScript
Примеры инструкций
Примеры SQL
Примеры Python
Примеры W3.